What is Dedifferentiation?
Under certain conditions, plants cells that are already differentiated and lost the ability of further division regain the capacity of division and differentiation. This process is known as dedifferentiation. The fully differentiated parenchyma cells undergo dedifferentiation, which leads to the formation of cork cambium and inter-fascicular cambium. A dedifferentiated tissue has the ability to act as meristem that could give rise to a different set of cells. The ability of those cells for further differentiation depends on different parameters such as genetic and epigenetic variations. This concept is used in plant tissue culture to develop a callus.
What is Redifferentiation?
Once new cells are formed from the dedifferentiated tissues that act as meristems, the cells lose their ability for further division and differentiation. Eventually, they get mature in order to accomplish specific functions of the plant body. Secondary xylem and secondary phloem are the best examples to describe the process of redifferentiation. Dedifferentiated vascular cambium divides further to give rise to the secondary xylem in the inside and secondary phloem on the outside. The secondary phloem and secondary xylem cells lose their ability for further division; instead, they become mature to fulfill specific functions of the plant body, which include transportation of food and water, respectively. Phelloderm is a layer of secondary tissues that is produced by the dedifferentiated cork cambium. Similar to secondary xylem and phloem, phelloderm’s cells lose their ability for further differentiation but become mature in order to perform specific functions such as limitation of dehydration and prevention of the entry of pathogens into the plant body due to the destruction of the epidermis.
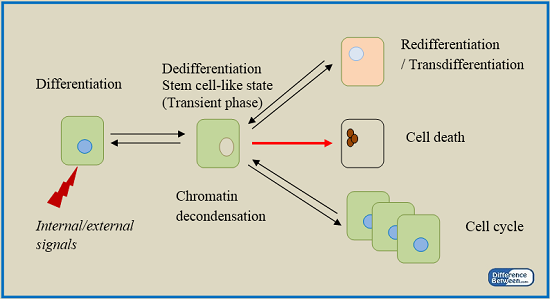
Figure 01: Differentiation and Redifferentiation



{kind=link}
{kind=link}